![]() Cell
Communication
Cell
Communication
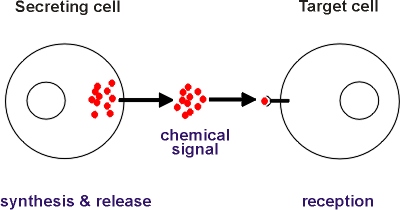
Cells communicate with each other by means of chemical signals (See
Figure). In multicellular organisms such communication is essential to
control development and to co-ordinate the diverse activities of cells
specialized to perform different functions such as secretion and contraction.
 |
Basic
Features of Cell Communication
Cells communicate with each other by means of chemical signals. The chemical signal is synthesised and released by the secreting cell, it is transported to a target cell where it is recognised and causes the taget cell to alter its activity in some way. The chemical signals can be pheromones, hormones, local hormones, neurotransmitters, cell surface molecules or intercellular messengers.
|
Cells have evolved chemical signalling pathways that enable them to communicate over long (pheromones and hormones), intermediate (local hormones) or very short (neurotransmitters, cell surface molecules, intercellular messengers) distances. These different chemical signals thus span a range of distances from nanometres right up to kilometres.
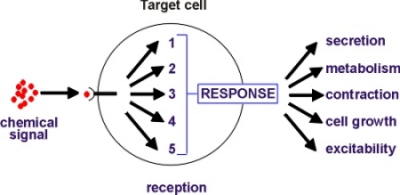
Target cells respond to chemical signals using a limited repertoire
of five intracellular signalling pathways that decode incoming signals
by processing them into forms that can be recognized by the response elements
that bring about the change in activity (see Figure).
|
Signal Reception and Processing
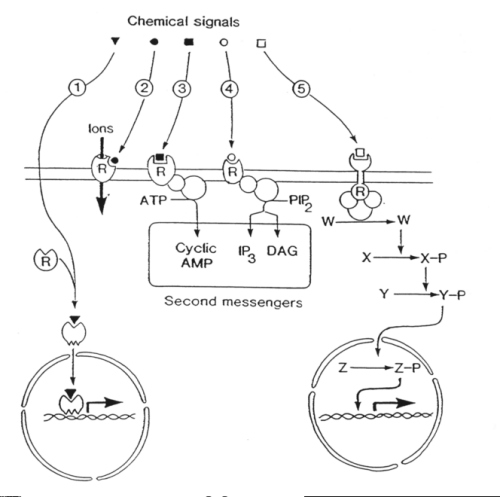
The five major signalling systems in cells all begin with a chemical signal binding to a receptor. One of these is used by the steroid hormones that bind to intracellular receptors whereas the other four pathways begin with hydrophilic chemical signals binding to plasma membrane receptors on the cell surface: |
 |
The five major intracellular signalling systems comprise:
1) Steroid hormone action.
Steroid hormones diffuse across the plasma membrane of the target cell
and bind reversibly to specific steroid hormone receptors in the cytosol.
The steroid-receptor complex then undergoes a conformational change which
allows it to bind to DNA to switch on the transcription of early response
genes which produce signals to activate other genes.
2) Ion channel-linked receptors.
Ion channel-linked receptors are used by neurotransmitters such as
acetylcholine. In the case of nicotinic receptors, acetylcholine binds
to sites on the outside to induce a conformational change which admits
a pulse of Na+ ions resulting in membrane depolarization. Some
receptors gate calcium ions (Ca2+) which acts as an intracellular
second messenger (see pathway 4, below).
3) Cyclic AMP signalling pathway.
A widely-used pathway containing the following steps: (1) chemical
signal binds to a 7 membrane spanning receptor to induce a conformational
change; (2) the III cytoplasmic loop of the receptor activates a G protein
(Gs); (3) G protein activates adenylate cyclase; (4) adenylate cyclase
uses ATP to synthesize the second messenger cyclic AMP; (5) cyclic AMP
acts through a protein kinase to induce a response; (6) in sense organs,
cyclic AMP opens a Na+ channel leading to a depolarizing receptor
potential to send information to the brain.
4) Phosphoinositide signalling pathway.
This complex signalling pathway plays a central role in regulating
many cellular processes. Its initial steps are similar to those in the
cyclic AMP signalling pathway (above) :
(1) Chemical signal binds to, and induces a conformational
change in, a 7 membrane spanning receptor.
(2) The III cytoplasmic loop of the receptor activates
a G protein (Gq).
(3) Gq stimulates an amplifier, phospholipase C (PLC).
(4) PLC acts on a minor membrane phospholipid phosphatidyl
inositol 4, 5-bisphosphate (PIP2) to give diacylglycerol (DAG)
and inositol 1, 4,5-trisphosphate (InsP3). This represents a
bifurcation in the signalling pathway because both DAG and InsP3
are second messengers.
(5) DAG acts within the plane of the lipid bilayer
to stimulate its effector protein kinase C (PKC).
(6) InsP3 diffuses into the cytosol where
it acts on ion channels to release Ca2+ stored up in the endoplasmic
reticulum.
(7) The onset of the Ca2+ signal is augmented
by a process of calcium-induced calcium release (CICR).
(8) Ca2+ can act through a protein kinase
or
(9) It can use specific binding proteins (calmodulin,
troponin C) to induce contraction in muscle cells, or
(10) It can act directly on ion channels to influence excitability.
Calcium spiking and calcium waves
The present of the positive feedback process of CICR (step 7 above) results in interesting spatial and temporal patterning of calcium signals. Calcium signals appear as discrete spikes whose frequency depends on the concentration of the external signal suggesting that cells are employing a frequency-modulated (FM) rather than an amplitude-modulated (AM) coding system. In many cells, each spike initiates at a discrete point and then spreads as a wave travelling at velocities of 50-100µm/s. These Ca2+ waves are particularly evident in eggs during fertilization.
5) Tyrosine kinase-linked receptors.
The signalling pathway activated by tyrosine kinase-linked receptors
is of crucial importance in the control of cell proliferation. Many growth
factors utilize this signalling system which is designed to transfer information
from the cell surface into the nucleus. Tyrosine kinase-linked receptors
are examples of membrane-spanning receptors that combine an agonist-binding
site on the outside with an enzymatic activity on the inside. The latter
functions as a transducer to relay information to different amplifiers
thereby spawning a number of signalling pathways. The most important of
these is a protein phosphorylation cascade that incorporates important
proto-oncogenes (ras and raf -1) and culminates in gene transcription.
These five signalling pathways are summarised in the Figure below:
| The five major signalling pathways
1. Steroid homones 2. Ion channel-linked receptors 3. cAMP pathway 4. Phosphoinositide pathway 5. Tyrosine kinase-linked receptors
|
 |
![]()
![]()