Neural Oscillations
Oscillatory activity in neural matter is ubiquitous and can be observed in single neurons, mass neural activity (electroencephalographic activity, EEG, or magnetoencephalographic, MEG, or local field potentials, LFP)1,2,3,4. Examples are shown below from sensorimotor EEG in man and primary motor cortex LFP in monkeys, where oscillations in the beta range (15-30Hz) are evident (underlined).
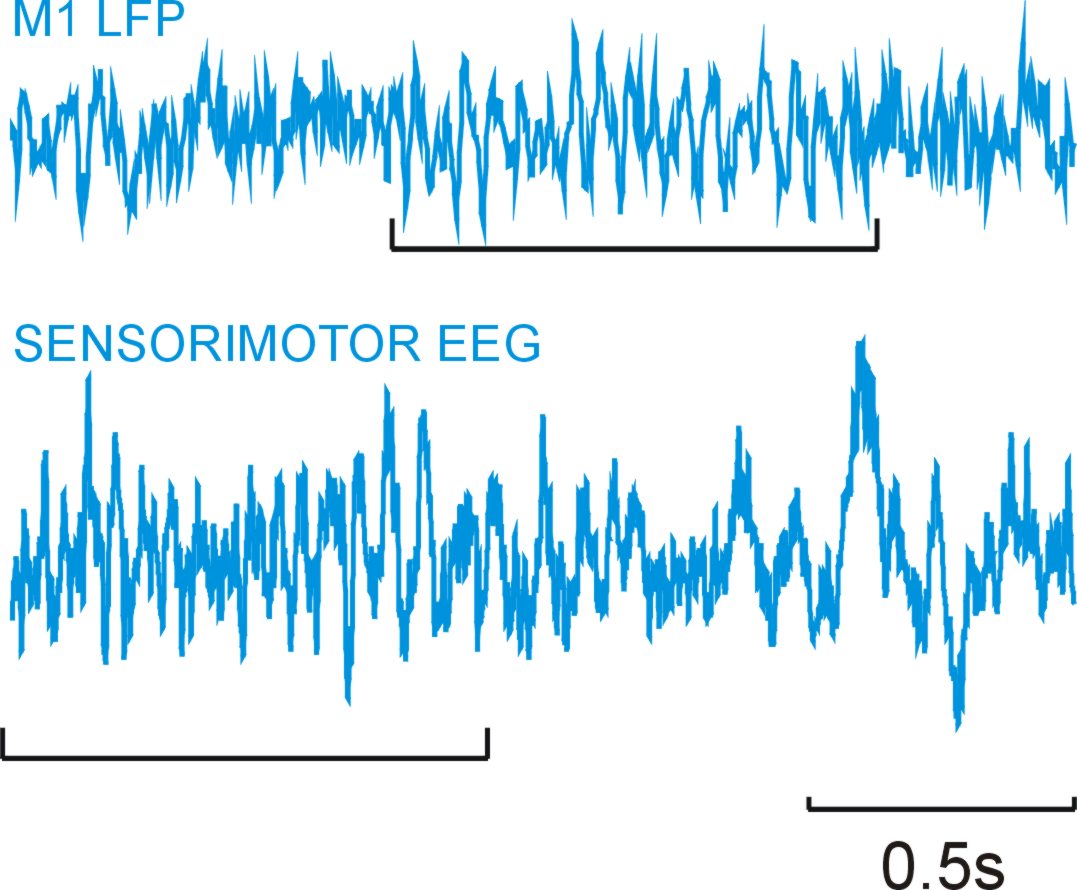
Oscillations can be observed from a variety of cortical areas, and can occur over a broad range of frequencies. In the motor system the most obvious oscillatory phenomena are around 8-40Hz. Do they have any functional significance, or are they simply an epiphenomenon in the same way that car engine noise is a by-product of combustion (and has no role in propulsion)?
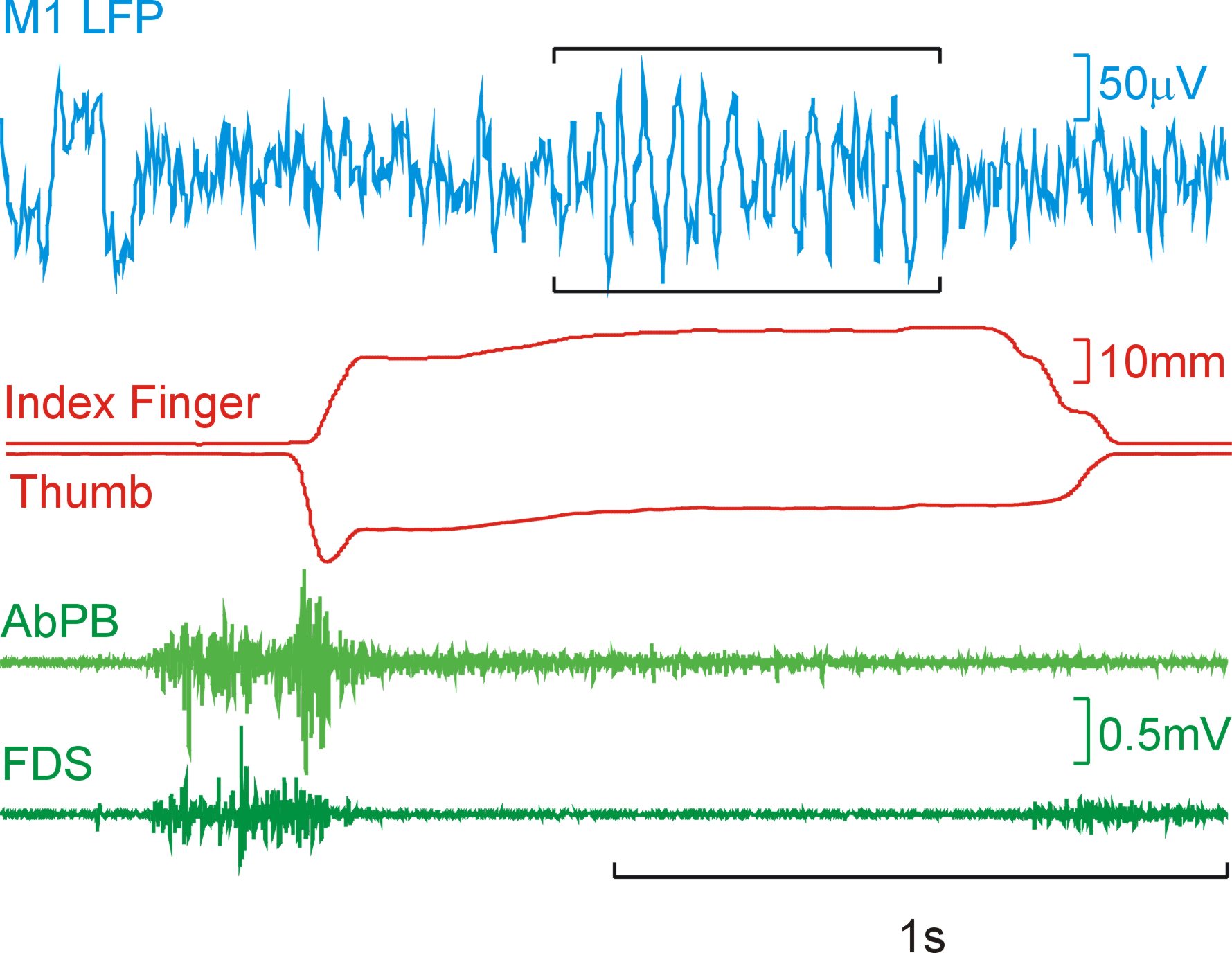
Regardless of their underlying functional role, their strength is modulated by behaviour. Below is a trace of LFP from M1 along with lever traces and muscle activity (EMG). As can be seen during the steady hold period there are visible oscillations in the LFP, but they disappear during periods of dynamic movement (when the levers are squeezed and released).
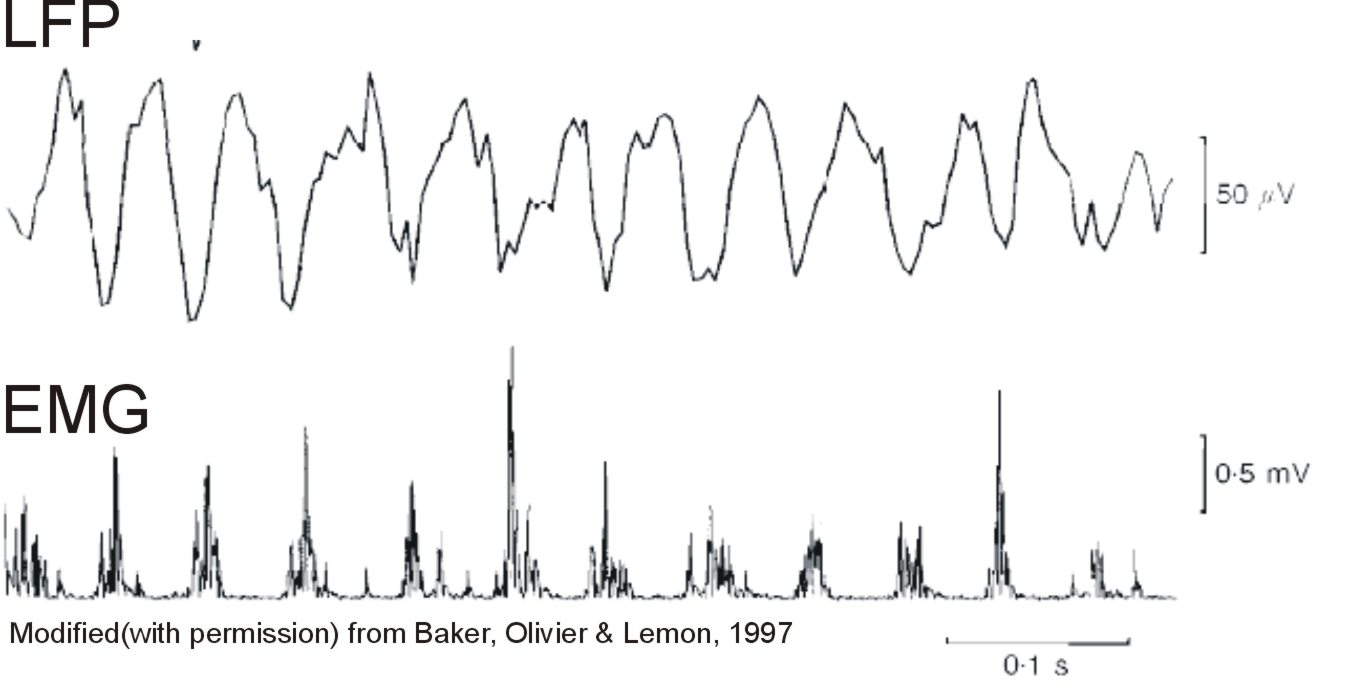
In some circumstances, during periods of steady contraction, coherence (correlation in the frequency domain) can be seen between oscillations in the cortex and muscle activity at ~20Hz, as shown by the figure below.
It has proven very hard to provide definitive evidence for a functional role of oscillations in the sensorimotor system specifically because they disappear during active movements. A recently proposed hypothesis suggests that these oscillations may be used to probe the state of the muscles right after a dynamic movement, with some supporting evidence5.
I have recorded from multiple single units in the deep cerebellar nuclei 6 (interpositus/dentate) during the performance of a precision grip task and have found that the cells in the cerebellum can phase lock with oscillations in M1 LFP.
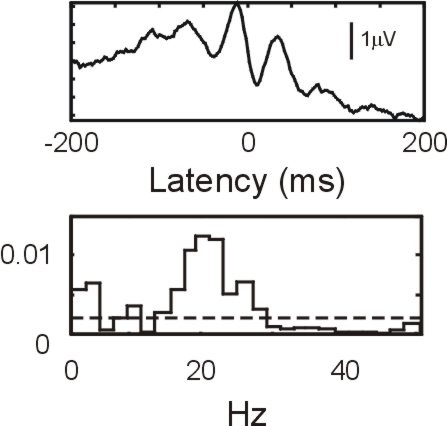
The above figure (top plot) shows the spike triggered average of an M1 LFP from a cerebellar output neurone, showing a clear oscillatory effect. The bottom plot is the frequency domain analogue of the spike triggered average, which shows that the significant (dotted line,P=5%) coherence between neural spiking and M1 LFP is ~15-30Hz. This suggests that the cerebellar output cells are weakly phase locked with oscillations in M1. As most of the cells in the deep cerebellar nuclei are output cells, these oscillations will be transmitted to their targets.
Even if neural oscillations are merely an epiphenomenon of some underlying neural process they are still of interest. To use the engine noise analogy again, even if the noise produced by the car engine is not used for actually moving the car, a loud clanking sound can still be useful in telling you that something has gone wrong!
- 1: Baker SN, Olivier E, Lemon RN. Coherent oscillations in monkey motor cortex and hand muscle EMG show task-dependent modulation. J Physiol. 1997 May 15;501 (Pt 1):225-41.
- 2: Conway BA, Halliday DM, Farmer SF, Shahani U, Maas P, Weir AI, Rosenberg JR. Synchronization between motor cortex and spinal motoneuronal pool during the performance of a maintained motor task in man. J Physiol. 1995 Dec 15;489 ( Pt 3):917-24.
- 3: Sanes JN, Donoghue JP. Oscillations in local field potentials of the primate motor cortex during voluntary movement. Proc Natl Acad Sci U S A. 1993 May 15;90(10):4470-4.
- 4: Murthy VN, Fetz EE. Oscillatory activity in sensorimotor cortex of awake monkeys: synchronization of local field potentials and relation to behavior. J Neurophysiol. 1996 Dec;76(6):3949-67.
- 5: Baker SN. Oscillatory interactions between sensorimotor cortex and the periphery. Curr Opin Neurobiol. 2007 Dec;17(6):649-55.
- 6: Soteropoulos DS, Baker SN. Cortico-cerebellar coherence during a precision grip task in the monkey. J Neurophysiol. 2006 Feb;95(2):1194-206.
Reference List