Bimanual Movements
Most of our everyday movements involve both hands to varying extents, be it to tie our shoe laces, to drive a car or to even open a bottle of water. It is clear that the two hands are not independent of each other, and movement of one has an impact on movement of the other 1,2 . A nice example is attempting to tap the head with one hand and to rub the tummy with the other; the result is that both hands tend to move together, each making a mixture of the intended tapping and rubbing movements.
A substantial amount is known about the behavioural constraints of bi-manual movements 1,2,3,4 , as well the activity of candidate brain areas, such as supplementary motor area (SMA), primary motor cortex (M1), reticular formation (RF) and even the cerebellum (Cb 5 ). However for any sort of co-ordination between the two hands, there needs to be communication between the systems controlling each hand. It is currently not clear what information is conveyed from one side of the brain to the other (see ref. 6), and how this is processed.
I have recorded from identified cells in the corpus callosum in SMA 6 during movement. These cells had very low baseline rates and phasic responses to hand movements (see image below) during a precision grip task with either left, right or both hands together.
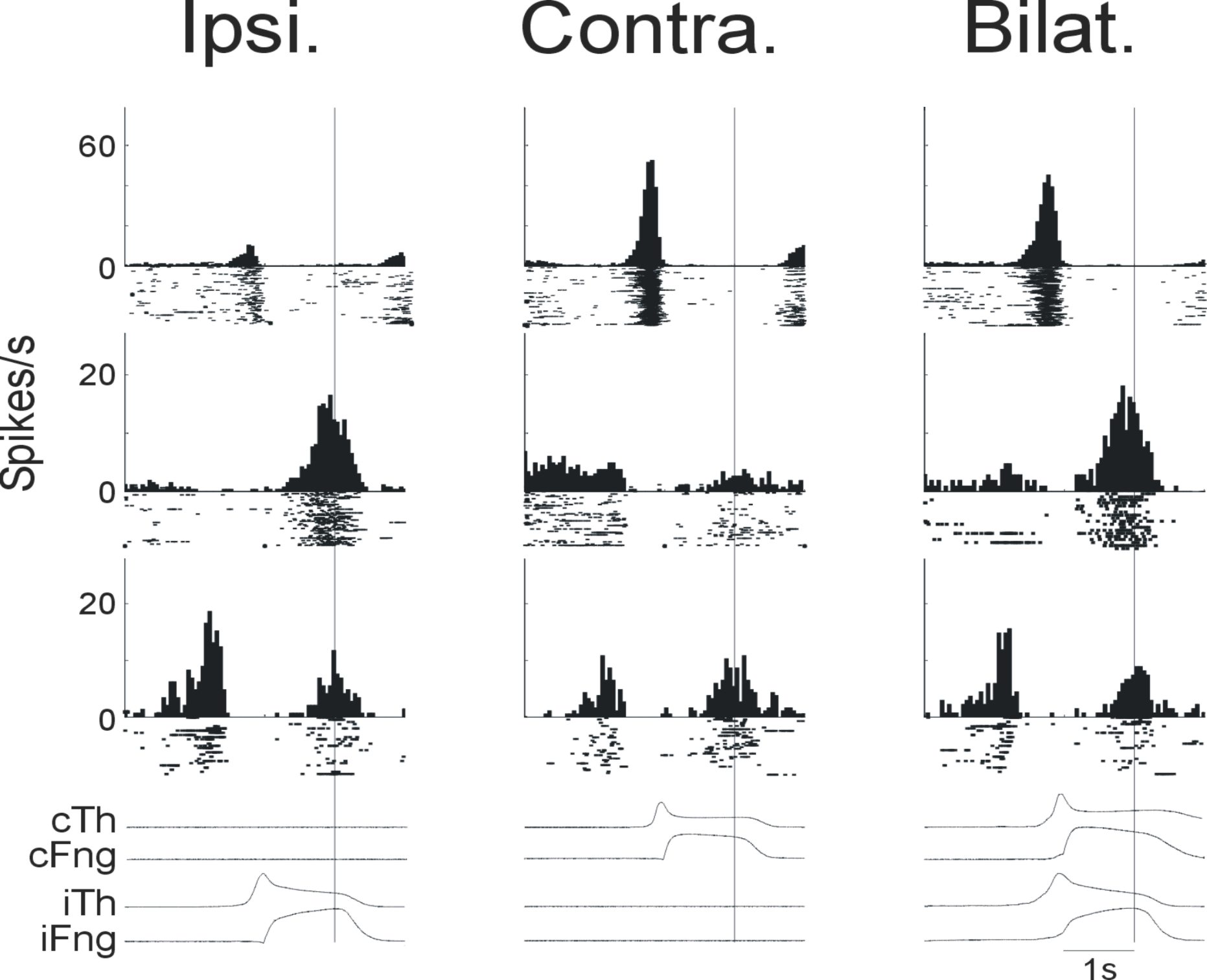
Each row above corresponds to a single callosal cell and each column corresponds to trial type (ipsilateral, contralateral or bilateral). The traces under the PSTH and rasters are the lever position traces for the thumb and finger.
Recording from identified callosal cells offers the luxury of interpreting their activity in a bimanual context regardless of the movement. Whatever activity these cells show it will be transmitted to the contralateral hemisphere. As their baseline activity is quite low, the callosal cells would be poorly suited to transmit information regarding a continuous signal, but instead their phasic responses make them much better suited to transmit information about timing i.e. 'when' something occurs.
In the same series of experiments I also recorded from, and stimulated, the output structures of the cerebellum, the deep cerebellar nuclei and found that they can show a certain degree of bilateral organisation 5, 6 . Some cells modulated their activity with movements of the contralateral arm, even though the cerebellum as classically considered an ipsilaterally organised structure. I also observed that low intensity stimulation (figure below) could produce effects in forearm and arm muscles bilaterally.
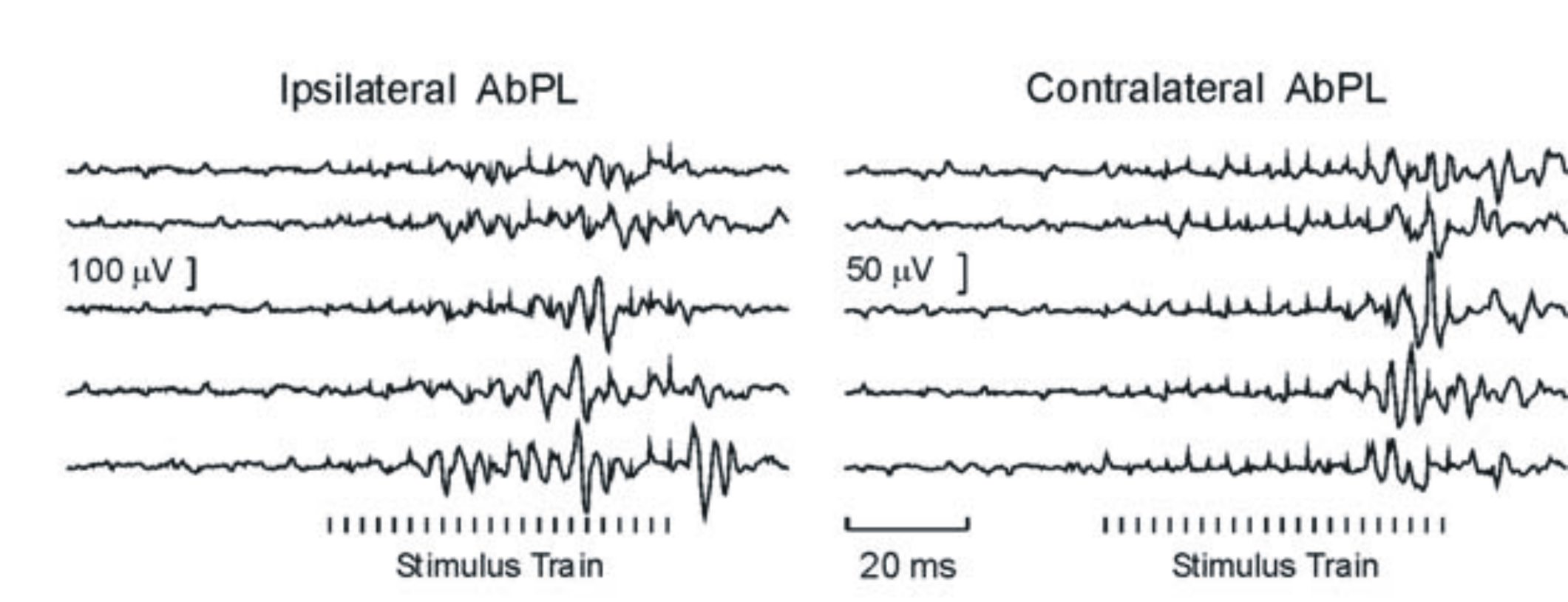
Single trial responses of Abductor Pollicis Longus activity to a stimulus train at 30microAmps
The pathway mediating these cerebellar effects is not clear at the moment, but one potential candidate is the brainstem reticular formation, as it is known to project to the spinal cord bilaterally and it receives cerebellar inputs.
- 1)Franz EA, Zelaznik HN, McCabe G (1991) Spatial topological constraints in a bimanual task. Acta Psychol (Amst) 77: 137-151.
- 2)Kelso JA, Southard DL, Goodman D. (1979) On the nature of human interlimb coordination. Science. 203:1029-31.
- 3) Kazennikov O, Perrig S, Wiesendanger M. (2002) Kinematics of a coordinated goal-directed bimanual task. Behav Brain Res. 134:83-91.
- 4) Perrig S, Kazennikov O, Wiesendanger M. (1999) Time structure of a goal-directed bimanual skill and its dependence on task constraints. Behav Brain Res. 103:95-104.
- 5) Soteropoulos DS, Baker SN (2008). Bilateral representation in the deep cerebellar nuclei. Journal of Physiology 586, 1117-1136.
- 6) Soteropoulos DS, Baker SN (2007). Different contributions of the corpus callosum and cerebellum to motor coordination in monkey. Journal of Neurophysiology 98, 2962-2973.
Some (of many) relevant references: