Neural Variability
Neurons carry information to other neurons by a sequence of action potentials, which are usually quite irregular (an example shown below).
The distribution of the times between spikes (interspike intervals, ISI) in real neurons often resembles a skewed distribution, with a long rightward tail corresponding to long ISI and a sharper leftward edge due to neural refractoriness (example of ISI histogram shown below).
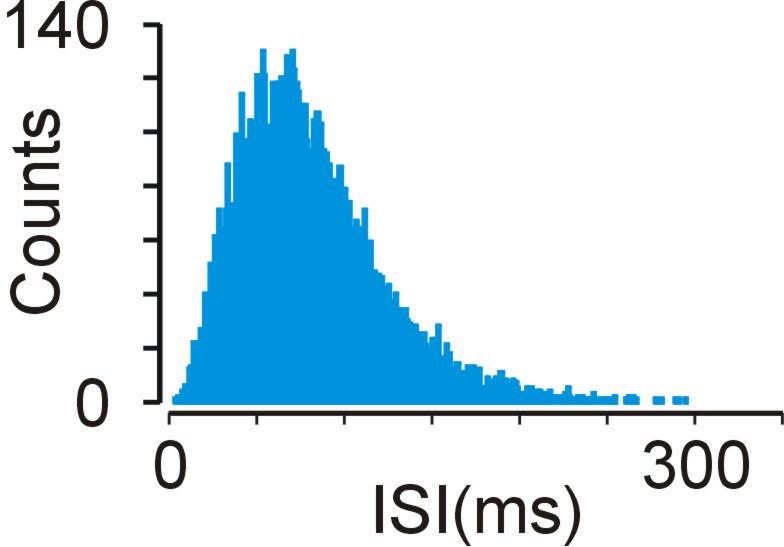
Is this variability simply noise (i.e. has nothing to do with the signal that the neuron is carrying) or is there some information in the specific temporal pattern of spiking? This has been debated for some time now and while there is some evidence to support the idea that there is information in the temporal patterns of spikes, there is no uniform consensus.
Neural activity related to a stimulus can vary in several other ways beyond the 'intrinsic' variability in spiking mentioned above. There can be variations in the latency of the neural response, the amplitude and even the shape of the response itself. The figure below shows the PeriStimulus Time Histogram (PSTH1) and raster plot of a simulated spike train with a rate step. There is a baseline rate of 30Hz followed by a rate increase to 80Hz for 300ms. The left PSTH & raster are for when there is no latency variability from trial to trial. However, when the single trial spike trains are jittered randomly (100ms stand. dev.), it becomes obvious (PSTH & raster on right) that the average response is a poor representation of what is going on in the single trials.
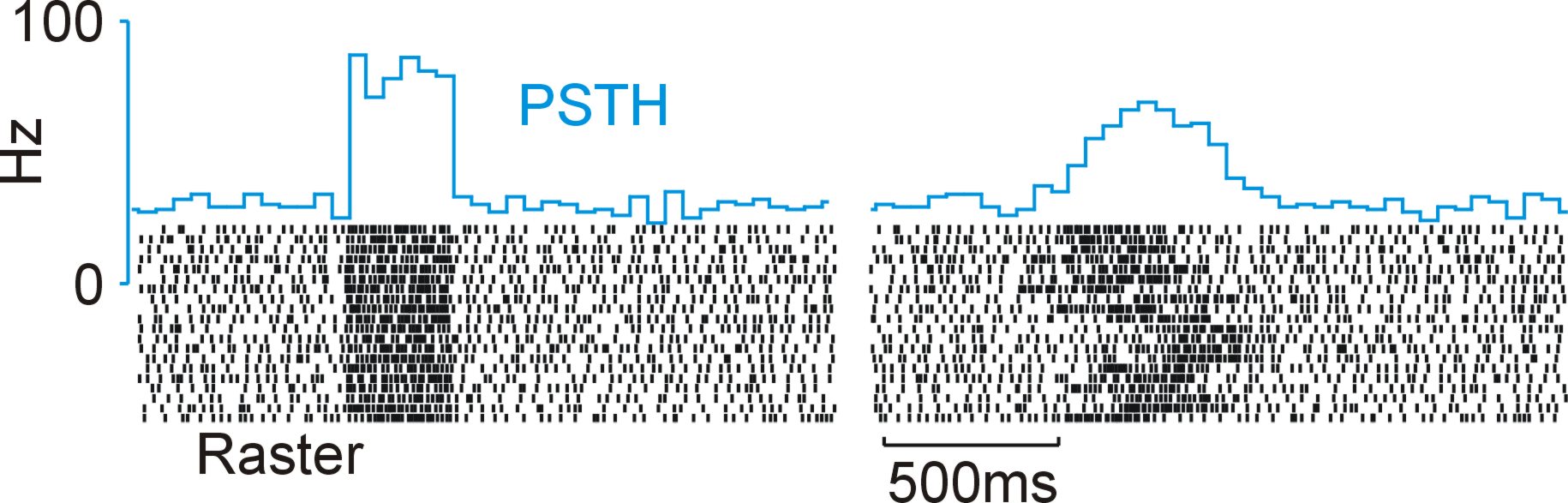
There are proposed methods that can mitigate the effects of such trial-to-trial variability or allow measurements from single trials 2,3,4,5
However, trial-to-trial variability is a physiological phenomenon that is interesting in its own right, and worth measuring 6,7,8 . Variability in response latency and/or amplitude may provide important clues to what a cell is coding for 9 . Highly reliable firing, time locked to a behavioral event, may signal the coding of that event. Greater variability may simply be noise, but more likely represents the encoding of another variable; this may not be experimentally observable (an internal “hidden state”) Trial-averaged measures such as the PSTH discard valuable information about the trial-to-trial response variability. An important question is what impact this trial-to-trial variability has in the information that single neurons can convey to their targets regarding the timing of an external event?
I have worked on a novel analysis method that measures this information10. The method proceeds by comparing single-trial responses with the “archetypal” response, estimated by the trial-averaged response in the PSTH. A cell is assigned an information value (in bits), representing how well it encodes the timing of a behavioral event or stimulus. This singular information value will be affected by a host of factors such as variability in response latency and response amplitude from trial to trial, as well as response size, baseline rate, and the intrinsic variability of neural spiking.
Reliable neurons, which have single-trial responses similar to the PSTH, return high information values. Rather than attempting to account for any latency or amplitude changes from trial to trial, these are treated as genuine features of the cell’s response because such variability degrades the cell’s ability to represent the time of the alignment event and leads to smaller information values. The method permits a novel interpretation of cell firing and can reveal which of several events a neuron codes best.
To find out more about this please read the paper10. To watch a presentation on this and other issues regarding spike trains and their analysis, please go to the Spike Train Analysis Network homepage. This Network is funded by the EPSRC to promote collaboration and the development of novel analysis techniques for neural data.
- 1: Gerstein GL, Kiang NY. An approach to the quantitative analysis of electrophysiological data from single neurons. Biophys J. 1960 Sep;1:15-28.
- 2: Baker SN, Gerstein GL. Determination of response latency and its application to normalization of cross-correlation measures. Neural Comput. 2001 Jun;13(6):1351-77.
- 3: Grźn S, Riehle A, Diesmann M. Effect of cross-trial nonstationarity on joint-spike events. Biol Cybern. 2003 May;88(5):335-51
- 4: Nawrot M, Aertsen A, Rotter S. Single-trial estimation of neuronal firing rates: from single-neuron spike trains to population activity. J Neurosci Methods. 1999 Dec 15;94(1):81-92.
- 5: Nawrot MP, Aertsen A, Rotter S. Elimination of response latency variability in neuronal spike trains. Biol Cybern. 2003 May;88(5):321-34.
- 6: Andolina IM, Jones HE, Wang W, Sillito AM. Corticothalamic feedback enhances stimulus response precision in the visual system. Proc Natl Acad Sci U S A. 2007 Jan 30;104(5):1685-90.
- 7: Gur M, Snodderly DM. High response reliability of neurons in primary visual cortex (V1) of alert, trained monkeys. Cereb Cortex. 2006 Jun;16(6):888-95.
- 8: Kara P, Reinagel P, Reid RC. Low response variability in simultaneously recorded retinal, thalamic, and cortical neurons. Neuron. 2000 Sep;27(3):635-46.
- 9: DiCarlo JJ, Maunsell JH. Using neuronal latency to determine sensory-motor processing pathways in reaction time tasks. J Neurophysiol. 2005 May;93(5):2974-86.
- 10: Soteropoulos DS, Baker SN. Quantifying neural coding of event timing. J Neurophysiol. 2009 Jan;101(1):402-17.
References